Jakieś 60 lat temu dwaj badacze, Joe Hin Tjio i Albert Levan, odkryli, że liczba chromosomów (kariotyp) u ludzi wynosi 46 chromosomów, czyli 23 pary, a nie 48, jak wcześniej sądzono (1). Kluczem do tego odkrycia było wprowadzenie serii ulepszeń w technikach hodowlanych stosowanych wobec ludzkich fibroblastów, szczególnie w zakresie traktowania ich kolchicyną, która przerywa podziały komórkowe na odpowiednim dla obserwacji chromosomów etapie. Jest to tzw. metafaza, podczas której chromosomy ulegają skurczeniu, dzięki czemu mogą być prawidłowo widoczne przez mikroskop. Co ciekawe, te same usprawnienia zostały wykorzystane w kolejnych latach do ustalenia, że nasi najbliżsi przodkowie (duże małpy człekokształtne, takie jak szympansy, bonobo, goryle czy orangutany) posiadali 48 chromosomów. Jak i kiedy nastąpiła ta zmiana w liczbie chromosomów? A przede wszystkim, jaką rolę odegrała ta różnica w powstaniu naszego gatunku? Najnowsze przełomy w technikach analizy genetycznej czynią ogromne postępy w rozwiązywaniu tych dwóch pytań.

Fuzja chromosomów: przyczyna różnic
Na początku lat 60. techniki cytogenetyczne nie były jeszcze przygotowane do badań porównawczych chromosomów w ogóle, a u naczelnych w szczególności. Techniki, które zostały udoskonalone w późniejszym czasie w celu uzyskania kolorowych pasm i wzorów międzypasmowych w chromosomach ujawniły, że nasz drugi największy chromosom jest wynikiem fuzji dwóch chromosomów należących do naszych najbliższych przodków ewolucyjnych.
Jednak dopiero po latach 70. dotarły techniki molekularne pozwalające na bezpośrednią analizę DNA na chromosomach, przed którymi dogłębna charakterystyka rearanżacji chromosomowej odróżniającej nas od małp człekokształtnych nie była możliwa. I tak, zauważono, że mniej więcej w centrum naszego chromosomu 2 znajdują się telomeryczne i subtelomeryczne sekwencje DNA (normalnie obecne tylko na jednym końcu chromosomów, ale nie w obszarach wewnętrznych) (2). Wynikało z tego jasno, że fuzja dwóch chromosomów była kompletna, to znaczy od jednego końca do drugiego. Obecnie, dostępność genomu człowieka i genomu małp człekokształtnych ujawniła, jak zawartość genetyczna naszego chromosomu 2 odpowiada sumie dwóch chromosomów naszych małpich przodków.
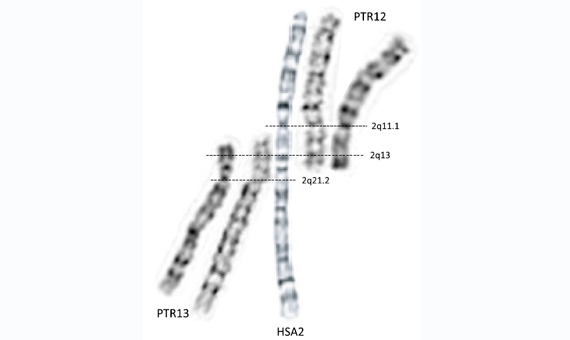
Znaleziono jednak również, że w obszarze fuzyjnym, który zapoczątkował nasz chromosom 2, brakuje pewnych regionów i sekwencji, które odpowiadają obszarom subtelomerycznym obecnym w dwóch chromosomach fuzyjnych u naszego gatunku. Innymi słowy, fuzja musiała wiązać się z utratą i rearanżacją części materiału genetycznego dwóch pierwotnie odrębnych chromosomów u przodków, których mamy wspólnych z małpami człekokształtnymi.
Denisowianie, neandertalczycy i małpy człekokształtne: kiedy się rozdzieliliśmy?
Analizy przeprowadzane obecnie na genomach wymarłych gatunków, które są z nami bezpośrednio spokrewnione, takich jak Denisowianie i Neandertalczycy, ujawniają, że gatunki te już przedstawiły fuzję chromosomów, która doprowadziła do powstania długiego chromosomu 2, charakterystycznego dla człowieka (3). Dlatego ta rearanżacja chromosomów sięga daleko w przeszłość: szacunki przy użyciu różnych metod datują to od 0,75 do 4,5 miliona lat temu.
Fakt, że Denisowianie i Neandertalczycy mieli tę samą liczbę chromosomów co my, może wyjaśniać, dlaczego potomkowie z międzygatunkowego krzyżowania z naszym gatunkiem byli zdolni do życia i prawdopodobnie płodni. Wyjaśniałoby to również, dlaczego ślady ich cech genetycznych pozostają w naszym genomie, jak wykazała porównawcza analiza genomiczna tych trzech gatunków. Jednak hipotetyczni potomkowie hodowli pomiędzy trzema wspomnianymi gatunkami hominidów (46 chromosomów) a ich przodkami małpami człekokształtnymi (48 chromosomów) mieliby problemy z niezgodnością chromosomową i prawdopodobnie nie byliby zdolni do życia. W rzeczywistości w naszym genomie nie znaleziono żadnych śladów specyficznych cech genetycznych małp człekokształtnych. Dlatego fuzja chromosomowa mogła zadziałać jako skuteczny mechanizm izolacji reprodukcyjnej, który odizolował nas od przodków małp człekokształtnych.
Na koniec, istnieje możliwość, że fuzja chromosomowa, z której powstał nasz chromosom 2, mogła być związana z pojawieniem się naszych charakterystycznych cech. Tak więc, kilka genów w naszym chromosomie 2, które znajdują się w pobliżu obszaru fuzji chromosomowej, ulega ekspresji intensywniej u naszego gatunku niż u małp człekokształtnych. Geny te ulegają ekspresji przede wszystkim w bardzo ważnych tkankach i organach, takich jak mózg i gonady (4). Po drugie, utrata pewnych sekwencji DNA, która nastąpiła w wyniku fuzji, mogła mieć „pozytywne” skutki dla naszych przodków.
Aby ostatecznie wyjaśnić, co nastąpiło w rearanżacji chromosomów, która jest tak charakterystyczna dla naszego gatunku, w nadchodzących latach będziemy musieli podjąć próbę uzyskania DNA wymarłych gatunków, które są starsze niż Homo Erectus czy Homo Heidelbergensis, określając w ten sposób, czy fuzja jest związana ze wszystkimi „ludzkimi” liniami; lub wykonując dogłębną analizę porównawczą obszaru fuzji naszego chromosomu 2 i subtelomerycznych obszarów dwóch dużych chromosomów małp biorących udział w fuzji, co do tej pory nie było możliwe.
Manuel Ruiz Rejón
Granada University, Autonomous University of Madrid
- J. Tjio i A. Levan. 1956. The chromosome number of Man. Hereditas, 42( 1-2): 1-6.
- W. Ijdo et al.1991. Origin of human chromosome 2: an ancestral telomere-telomere fusión. PNAS, 88: 9051-9056.
- Meyer et al. 2012 A high-coverage genome sequence from an archaic Denisovan individual. Science, 338:222-226; K. H. Miga. 2016. Chromosome-specific Centromere sequences provide an estímate of the Ancestral Chromosome 2 Fusion event in Hominin Genome.Journ. of Heredity. 1-8. Doi:10.1093/jhered/esw039.
- GTEx Portal, http://www.gtexportal.org/home.
.