 br>>>/div>
br>>>/div>Um Polvo Gigante do Pacífico Norte (Enteroctopus dofleini) vive de três a quatro anos; põe milhares de ovos numa única vez e depois morre. Em contraste, uma árvore de pau-brasil maduro (Sequoia sempervirens) vive durante muitas centenas de anos e produz milhões de sementes todos os anos (Figura 1). Como estes dois exemplos ilustram, os organismos diferem dramaticamente na forma como se desenvolvem, no tempo que levam a crescer, quando se tornam maduros, quantos descendentes de um determinado tamanho produzem, e quanto tempo vivem. Em conjunto, os padrões de desenvolvimento, crescimento, maturação, reprodução, sobrevivência, e duração de vida de um organismo definem o seu ciclo de vida, a sua história de vida.
 br>>>div>
br>>>div>
O principal objectivo da teoria da história de vida, um ramo da ecologia evolutiva, é explicar a notável diversidade das histórias de vida entre as espécies. Mas há outra razão mais convincente para a importância da evolução da história de vida: a adaptação por selecção natural baseia-se na variação da aptidão física darwiniana entre indivíduos, e uma vez que os traços da história de vida determinam a sobrevivência e a reprodução, eles são os principais componentes da aptidão física. O estudo da evolução da história de vida trata, portanto, de compreender a adaptação, a questão mais fundamental da biologia evolutiva.
p>Aqui apresentamos os fundamentos da teoria da história de vida e revemos o que os biólogos aprenderam sobre a evolução da história de vida. Para uma cobertura mais aprofundada, referimo-nos a Stearns (1992), Roff (1992, 2002), Charlesworth (1994), e Flatt e Heyland (2011). Ver também os artigos Nature Education Knowledge de Shefferson (2010), Young (2010), e Fabian e Flatt (2011).
The Basics of Life History Theory
Teoria da história da vida procura explicar como a selecção natural e outras forças evolutivas moldam os organismos para optimizar a sua sobrevivência e reprodução face aos desafios ecológicos colocados pelo ambiente (Stearns 1992, Roff 1992, Stearns 2000), ou como David Reznick o disse recentemente: “A teoria da história da vida prevê a forma como a selecção natural deve moldar a forma como os organismos parcelam os seus recursos para fazer bebés” (Reznick 2010, p. 124). A teoria fá-lo analisando a evolução dos componentes da aptidão física, os chamados traços de história de vida, e como interagem: tamanho à nascença; padrão de crescimento; idade e tamanho à maturidade; número, tamanho e sexo da prole; esforço reprodutivo específico da idade, fase ou tamanho; taxas de sobrevivência da idade, fase ou tamanho específico; e duração da vida.
A teoria clássica trata a evolução da história de vida como um problema de optimização: dados os factores ecológicos particulares (por exemplo, predadores, nutrição) que afectam a probabilidade de sobrevivência e reprodução de um organismo, e dados os constrangimentos limitantes e os trade-offs intrínsecos ao organismo, quais são os valores óptimos e as combinações de traços da história de vida que maximizam o sucesso reprodutivo? Para encontrar a solução para este problema, precisamos de compreender as suas “condições limite” (Stearns 2000): (1) como factores extrínsecos e ambientais afectam a sobrevivência e a reprodução; e (2) como as ligações intrínsecas entre os traços da história de vida (trade-offs) e outras limitações limitam se e como os traços da história de vida podem evoluir. Uma vez que estas condições tenham sido compreendidas e definidas, os modelos de história de vida podem ser utilizados para responder a perguntas como, por exemplo: Quão pequeno ou grande deve um organismo crescer? Com que idade e tamanho deve ele amadurecer? Quantas vezes deve reproduzir-se? Quantos descendentes deve produzir e de que tamanho devem ser? Durante quanto tempo deve reproduzir-se e quanto tempo deve viver?
Problemas de optimização do histórico de vida são tipicamente modelados usando a equação de Euler-Lotka, que descreve a taxa de crescimento populacional (ou seja aptidão) de um genótipo clonal (ou substituição de alelos) em tempo de geração contínua em função da sua idade na maturidade, idade na última reprodução, probabilidade de sobrevivência específica da idade, e o número esperado de descendência específica da idade (Stearns 1992, Roff, 1992, Brommer 2000):
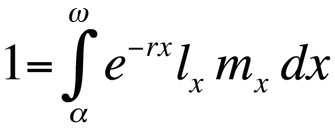
onde α é a idade na primeira reprodução, ω a idade na última reprodução, lx a probabilidade de sobrevivência desde o nascimento até à classe de idade x, mx o número esperado de descendentes na classe de idade x, e r a taxa de crescimento populacional ou parâmetro Malthusiano. A equação soma as probabilidades de sobrevivência e reprodução ao longo de toda a vida dos indivíduos da população e pode então ser resolvida para r. Note-se que no contexto da teoria da história de vida r mede a taxa de crescimento ou aptidão de um clone ou, em organismos sexualmente reprodutores, a taxa de propagação de um alelo que afecta a história de vida. Assim, a hipótese implícita é que a população modelada é constituída por indivíduos fenotípica e geneticamente idênticos. Se a população descrita pela equação de Euler-Lotka for estacionária (sem crescimento), r é zero e a equação torna-se
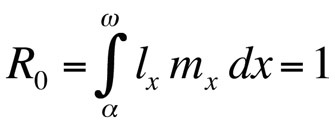
ou, se o tempo de geração for discreto,
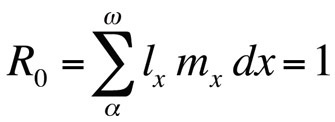
onde R0 é o número esperado de filhas por fêmea por vida (taxa reprodutiva líquida). Esta equação é mais simples que a versão de tempo contínuo e pode ser usada sempre que r é zero ou próximo de zero; para populações estáveis que não mudam de tamanho, R0 é a medida de aptidão adequada (Stearns 1992, Roff 1992, Brommer 2000). Usando esta estrutura, pode-se perguntar que combinação particular de traços de história de vida maximiza a aptidão física, ou quanto a aptidão física é afectada quando um dos traços é alterado. Esta abordagem tem sido utilizada com grande sucesso para prever a evolução dos traços da história de vida.
Variação genética dos traços do historial de vida
A evolução dos traços do historial de vida por selecção natural depende da variação genética sobre a qual a selecção pode actuar para produzir adaptações em resposta ao ambiente. Os modelos acima mencionados assumem implicitamente que a evolução da história de vida não está limitada por uma falta de variação genética. Curiosamente, contudo, a hereditariedade (h2 = VA/VP = variação genética aditiva dividida pela variação fenotípica), ou seja, a proporção de diferenças fenotípicas entre indivíduos de uma população que é explicada pelas diferenças genéticas aditivas entre eles, é normalmente pequena para os traços de história de vida. Esta baixa hereditariedade pode ser causada por baixas quantidades de variância genética aditiva; no entanto, existe uma ampla variação genética para traços de história de vida em populações naturais e laboratoriais. Consistentes com a noção de que os componentes de aptidão física abrigam muita variação genética, muitas experiências de selecção artificial no laboratório conseguiram causar com sucesso mudanças evolutivas nos traços de história de vida na direcção prevista (Stearns 1992, Roff 1992, Houle 2001). Uma razão para os grandes traços VA da história de vida pode ser o facto de serem traços altamente complexos, quantitativos e poligénicos influenciados por muitos loci (Houle 1992).
Mas como podemos conciliar o facto de que a VA é grande enquanto ao mesmo tempo a h2 é pequena? Uma razão provável para a baixa hereditariedade dos traços da história de vida é que, embora VA (o numerador) seja grande, VP (o denominador) é muito maior do que VA. Note-se que a variância fenotípica VP consiste em VA, a variância genética aditiva, mais um resto, VR, que por sua vez consiste em todas as fontes genéticas não aditivas de variação (isto é, devido a dominância, epistasia, etc.) e variação fenotípica engendrada pelo ambiente (isto é, plasticidade fenotípica, genótipo por interacções ambientais; ver abaixo). Assim, os traços da história de vida têm provavelmente baixa hereditariedade porque são influenciados por muitos loci (que infla tanto VA como VP) ao mesmo tempo que abrigam quantidades substanciais de variação residual VR, por exemplo variação devido a mudanças no ambiente (que inflaciona VP mas não VA) (Houle 1992, Houle 2001).
Mais, embora os traços de história de vida estejam sob forte selecção, o que deveria esgotar a variância genética, vários factores podem manter a variação genética para estes traços, incluindo o equilíbrio entre mutação-selecção, heterogeneidade ambiental e genótipo por interacções ambientais, e correlações genéticas negativas (Stearns 1992, Roff 1992, Houle 2001). No entanto, apesar da variação típica de grandes quantidades de história de vida, a evolução da história de vida está também sujeita a restrições.
Troca da história de vida e outras restrições
Fitness seria obviamente máxima se a sobrevivência e reprodução fossem máximas em todas as idades, fases, ou tamanhos de um organismo. Em princípio, então, o problema básico da evolução da história de vida é trivial: todos os traços da história de vida devem sempre evoluir de modo a maximizar a sobrevivência e reprodução e assim a aptidão física (Houle 2001). Isto levaria muito rapidamente à evolução dos “demónios darwinianos” (Lei 1979) que se apoderariam do mundo, ou seja, organismos que começam a reproduzir-se assim que nascem, produzem um número infinito de descendência, e vivem para sempre. Tais organismos, contudo, não existem no mundo real: Os recursos são finitos, e os traços da história de vida estão sujeitos a trocas intrínsecas e outros tipos de restrições, pelo que a selecção natural não pode maximizar os traços da história de vida – e portanto a aptidão – para além de certos limites. Chamamos a tais limites restrições evolutivas (Stearns 1992, Houle 2001); como mencionado acima, representam a “condição intrínseca de limite” que temos de compreender para prever a evolução da história de vida.
Um dos tipos mais importantes de restrições são os trade-offs da história de vida (Stearns 1992, Roff 1992, Flatt e Heyland 2011). Existe um trade-off quando um aumento num traço de história de vida (melhoria da aptidão física) é associado a uma diminuição noutro traço de história de vida (redução da aptidão física), para que o benefício da aptidão física através do aumento do traço 1 seja equilibrado contra um custo da aptidão física através da diminuição do traço 2 (Figura 2A). (Note-se que os trade-offs podem também envolver mais de dois traços.) A nível genético, pensa-se que tais trade-offs são causados por alelos com efeitos pleiotrópicos antagónicos ou por desequilíbrio de ligação entre loci.
Trade-offs são tipicamente descritos por correlações fenotípicas ou genéticas negativas entre os componentes da aptidão entre indivíduos de uma população (Figura 2A). Se a relação for genética, prevê-se uma correlação genética negativa para limitar (ou seja, para abrandar ou prevenir) a evolução dos traços envolvidos. Assim, existe um trade-off genético numa população quando uma mudança evolutiva num traço que aumenta a aptidão física está ligada a uma mudança evolutiva num outro traço que diminui a aptidão física. A existência de correlações genéticas pode ser estabelecida através de desenhos de reprodução genética quantitativa ou através de respostas fenotípicas correlacionadas à selecção. Por exemplo, a selecção artificial directa para uma maior longevidade em populações de laboratório geneticamente variáveis de moscas da fruta (Drosophila melanogaster) provoca a evolução do aumento da longevidade adulta (por vezes em 10 ou menos gerações), mas este aumento evolutivo da longevidade é associado a uma diminuição da reprodução precoce (por exemplo, Zwaan et al. 1995). Isto sugere que a longevidade e a reprodução precoce estão geneticamente associadas negativamente, por exemplo através de alelos pleiotrópicos antagónicos (por exemplo, Flatt 2011, Fabian e Flatt 2011).
Ao nível fisiológico, as compensações são causadas pela atribuição competitiva de recursos limitados a uma característica da história de vida em relação a outra dentro de um único indivíduo, por exemplo, quando indivíduos com maior esforço reprodutivo têm uma longevidade mais curta ou vice-versa (Figura 2B). Uma forma útil de pensar os trade-offs de atribuição de recursos é imaginar um historial de vida como sendo uma tarte finita, com as diferentes fatias representando a forma como um organismo divide os seus recursos entre crescimento, armazenamento, manutenção, sobrevivência e reprodução (Reznick 2010; Figura 2C). O problema essencial é este: dadas as circunstâncias ecológicas, e o facto de fazer uma fatia maior significa fazer outra mais pequena, qual é a melhor maneira de dividir a torta? Note-se que, uma vez que os trade-offs de atribuição de recursos podem ter uma base genética, e uma vez que diferentes genótipos podem diferir em aspectos da atribuição de recursos, as visões genética e fisiológica dos trade-offs não são necessariamente incompatíveis. Contudo, os trade-offs fisiológicos a nível individual nem sempre se traduzem em trade-offs genéticos (evolutivos) a nível da população. Por exemplo, quando o trade-off fisiológico (dentro do indivíduo) é geneticamente fixo (“hard-wired”) entre todos os indivíduos da população, todos os indivíduos exibirão a mesma relação fisiológica negativa entre dois traços da história de vida, mas a correlação genética entre indivíduos seria zero (Stearns 1989, Stearns 1992).
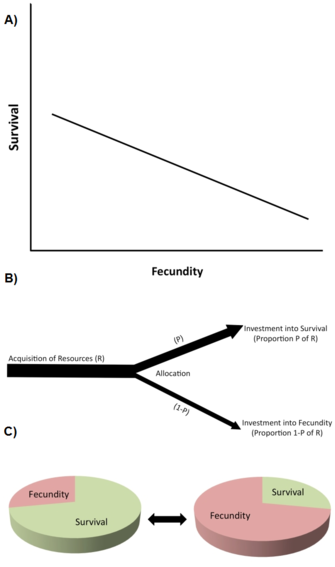
O livro de Stearns (1992) lista 45 possíveis trade-offs entre os 10 principais traços da história de vida, e muitos mais podem ser previstos para existir. Os trade-offs que têm recebido mais atenção incluem (1) reprodução actual versus sobrevivência; (2) reprodução actual versus reprodução futura; (3) reprodução actual versus crescimento parental; (4) reprodução actual versus condição parental; e (5) número versus tamanho da descendência.
Uma das melhores provas de trade-offs de história de vida de base genética vem da selecção artificial e experiências de evolução experimental realizadas em Drosophila (ver revisões em Stearns and Partridge 2001, Flatt and Schmidt 2009, Flatt 2011). Em resumo, muitas experiências encontraram: uma correlação negativa entre a fecundidade precoce e a vida adulta; uma correlação positiva entre o tempo de desenvolvimento e o tamanho do corpo; uma correlação positiva entre o tempo de desenvolvimento ou o tamanho do corpo com a fecundidade precoce; e uma correlação negativa entre a fecundidade precoce e a fecundidade tardia.
Outros constrangimentos nas histórias de vida que impedem a selecção natural de atingir uma determinada aptidão óptima podem envolver factores biofísicos, bioquímicos e estruturais, propriedades de desenvolvimento do organismo, contingências filogenéticas e históricas, ou simplesmente uma falta de variação genética (Stearns 1992, Houle 2001).
Plasticidade fenotípica nos traços da história de vida
A variação e constrangimentos genéticos não são os únicos factores que afectam a expressão e evolução dos traços da história de vida. Outra questão importante é que a variação do historial de vida é frequentemente fortemente influenciada pelo ambiente (por exemplo, temperatura, nutrição, predadores,), um fenómeno chamado plasticidade fenotípica, ou seja, a capacidade de um único genótipo (ou clone) produzir diferentes fenótipos em diferentes ambientes (Stearns 1992, Roff 1997, Pigliucci 2001, DeWitt e Scheiner 2004). A plasticidade de um genótipo específico pode ser conceptualmente descrita por uma função matemática chamada norma de reacção, ou seja, uma linha ou curva que relaciona os fenótipos produzidos por este genótipo com mudanças no ambiente que experimenta (Figura 3A).
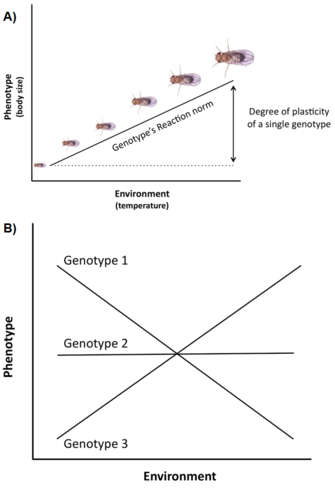
A importância de tal plasticidade na evolução da história da vida é pelo menos tripla (Stearns e Koella 1986, Stearns et al. 1991, Stearns 1992, Nylin e Gotthard 1998, DeWitt e Scheiner 2004, Flatt 2005): (1) uma vez que a plasticidade modula a expressão fenotípica da variação genética para traços únicos da história de vida e de correlações genéticas para pares de traços, afecta a resposta genética à selecção entre ambientes; (2) se existir variação adaptativa entre genótipos para a resposta plástica, a selecção pode produzir uma norma de reacção óptima que maximiza a aptidão física entre ambientes; e (3) a plasticidade de um traço específico pode proteger homeostaticamente o organismo contra alterações de outros traços induzidas pelo ambiente, de modo a optimizar o desempenho do organismo e, assim, a aptidão física.
Muitos traços de história de vida (por exemplo idade na maturidade, fecundidade) exibem um elevado grau de plasticidade, e existe frequentemente uma variação genética significativa de plasticidade em populações naturais, ou seja, os genótipos têm normas de reacção diferentes, um fenómeno chamado genótipo por interacção ambiental (GxE) (Figura 3B). Além disso, não só os traços individuais mas também as correlações entre traços podem ser plásticos, e diferentes ambientes podem alterar a inclinação e/ou o sinal da correlação de traços (Stearns et al. 1991, Stearns 1992). Nos sapos spadefoot (Scaphiophus couchii), por exemplo, os indivíduos que se desenvolvem em lagos de curta duração têm um período larval mais curto e um tamanho corporal menor na metamorfose (com os traços a serem negativamente correlacionados) do que os indivíduos que se desenvolvem em lagos de longa duração (com os traços a serem positivamente correlacionados) (Newman 1988, Stearns et al. 1991).
Having discutiu a abordagem de modelização da optimização e os factores que influenciam a expressão dos traços da história de vida, voltamo-nos agora para a discussão de algumas previsões importantes para a evolução das histórias de vida (para detalhes ver Stearns 1992, Roff 1992, Charlesworth 1994, Stearns 2000, Roff 2002).
Previsões para a evolução dos traços da história de vida
Em que idade e tamanho deve um organismo amadurecer? O sucesso reprodutivo de um genótipo depende fortemente da sua taxa de crescimento e – como consequência do crescimento – da sua idade e tamanho na maturidade. Para prever a idade e tamanho óptimos na maturidade, temos de compreender os custos e benefícios relativos (em termos de mortalidade e reprodução) quer da maturação precoce ou tardia, quer do crescimento em grande escala ou da permanência em pequena escala. Os benefícios de uma “estratégia” são os custos da outra, e vice-versa. Os benefícios de amadurecer mais cedo e com um tamanho menor (ou seja, os custos de amadurecer mais tarde e com um tamanho maior) incluem: (1) uma maior probabilidade de sobrevivência até à maturidade devido a uma duração mais curta do período de desenvolvimento arriscado e juvenil, e (2) um tempo de geração mais curto que permite aos pais produzir descendentes que nascem mais cedo e que começam a reproduzir-se mais cedo. Assim, a elevada mortalidade juvenil, por exemplo, deve favorecer a evolução de uma maturidade mais precoce. Inversamente, os benefícios de amadurecer mais tarde e com um tamanho maior (ou seja, os custos de amadurecer mais cedo e com um tamanho menor) incluem: (1) crescimento mais longo que leva a um maior tamanho na maturidade e, portanto, a um aumento da fecundidade (uma vez que a fecundidade aumenta frequentemente com o tamanho), (2) menor mortalidade adulta (e, portanto, uma fecundidade potencialmente maior durante a vida) devido a um maior tamanho (a mortalidade devida a predadores é frequentemente menor para indivíduos maiores), e (3) descendentes de maior qualidade (por exemplo, maior investimento por descendência, melhores cuidados parentais) que melhoram a sobrevivência da descendência produzida.
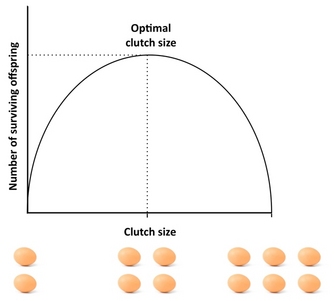
Quantos descendentes devem ser produzidos por um organismo? Um bom ponto de partida para abordar esta questão é a “falta de embraiagem”, um conceito que remonta ao ornitólogo David Lack (1947). A percepção básica de Lack era que as aves deveriam optimizar o tamanho da sua ninhada maximizando o número de descendentes (sobreviventes) de pombos. Desvios deste tamanho óptimo da ninhada reduziriam o sucesso reprodutivo: se os pais produzirem uma ninhada demasiado grande, poderão não ser capazes de suportar e criar toda a sua descendência, com alguns ou todos a morrer, enquanto que se produzirem uma ninhada demasiado pequena, o número de descendentes de pombos poderá ser inferior ao que os pais poderiam suportar (Figura 4). Embora Lack estivesse correcto na sua afirmação de que a aptidão física é frequentemente maximizada no investimento intermédio, os tamanhos da embraiagem são tipicamente menores do que o que a embraiagem Lack prevê. Isto porque o conceito ignora vários factores que podem reduzir o tamanho da embraiagem, incluindo a mortalidade dos pais, a reprodução futura, a taxa de produção dos netos, e o conflito entre pais e filhos. Por exemplo, a existência de trade-offs entre reprodução actual e sobrevivência dos pais, reprodução actual e futura, ou o número e tamanho da descendência pode causar desvios da Embraiagem Falta, e os modelos que levam tais factores em conta costumam produzir um melhor ajuste ao tamanho observado da embraiagem do que a Embraiagem Falta. Para um excelente estudo empírico do tamanho óptimo da embraiagem, ver o estudo de Daan et al. (1990) sobre peneireiros europeus.
Quantas vezes deve um organismo reproduzir-se? Deve ser semelparo (ou seja, reproduzir-se apenas uma vez) ou iteroparoso (ou seja, reproduzir-se várias vezes) (Figura 5; ver também Young 2010)? A teoria sugere que a iteroparidade e a evolução da vida reprodutiva aumentada são favorecidas quando a sobrevivência adulta é elevada e quando a fecundidade adulta ou a sobrevivência juvenil é baixa: a sobrevivência adulta elevada (ou não variável) aumenta o número de eventos reprodutivos por vida, e a fecundidade baixa (ou variável) ou a sobrevivência juvenil causam perdas de aptidão física que seleccionam para um maior esforço reprodutivo compensatório. Inversamente, a semelparidade e a evolução da diminuição da vida reprodutiva são favorecidas quando a sobrevivência adulta é baixa e a sobrevivência juvenil é alta. Assim, um adulto elevado (ou não variável) em relação à sobrevivência juvenil favorece a iteroparidade e prolonga a vida reprodutiva, enquanto que um adulto elevado (ou variável) em relação à mortalidade juvenil favorece a semelparidade e diminui a vida reprodutiva. Os organismos semelparos têm tipicamente um esforço reprodutivo mais elevado do que os organismos iteropários.

As considerações acima são aspectos de uma questão mais geral chamada “problema geral de história de vida” ou “modelo de esforço reprodutivo” (Schaffer 1983, Stearns 1992, Roff 1992, Charlesworth 1994): dado que a reprodução tem ambos os benefícios (i.e, produção da prole) e custos (por exemplo, diminuição da reprodução futura, aumento da mortalidade parental ou da prole), qual é o investimento reprodutivo óptimo que maximiza a aptidão física? Muitos estudos teóricos e empíricos têm abordado este problema; algumas das principais conclusões deste trabalho são (ver Stearns 1992, Roff 1992, Charlesworth 1994): (1) o esforço reprodutivo aumenta frequentemente, mas nem sempre, com a idade porque se espera que o custo da reprodução actual em termos de reprodução futura diminua com a idade (uma vez que o número de eventos reprodutivos futuros em que os custos se podem manifestar diminui com a idade); (2) se o esforço reprodutivo produzir rendimentos decrescentes ou se a mortalidade aumentar à medida que o esforço aumenta, o investimento reprodutivo intermédio e a iteroparidade são favorecidos; caso contrário, o esforço reprodutivo máximo e a semelparidade são favorecidos; (3) se a mortalidade aumentar em todas as classes etárias, o esforço reprodutivo aumenta cedo na vida e a idade na maturidade diminui; do mesmo modo, se a mortalidade adulta aumentar, a idade na maturidade deve diminuir; e (4) se a mortalidade aumentar após uma determinada idade (ou numa classe etária específica), o esforço reprodutivo aumenta antes e diminui depois dessa idade. Como testes particularmente belos de tais modelos de esforço reprodutivo, remetemos o leitor para as experiências de campo e laboratoriais de David Reznick e colegas em Trinidad guppies (por exemplo, Reznick et al. 1990).
Quanto tempo deve um organismo viver (ver também Fabian e Flatt 2011)? A evolução da esperança de vida pode ser vista como um equilíbrio entre a selecção para o aumento da esperança de vida reprodutiva (e, por conseguinte, potencial aumento do sucesso reprodutivo) e o envelhecimento (ou seja, aumento intrínseco da mortalidade dependente da idade). Os benefícios da evolução de uma vida reprodutiva mais longa incluem (ver também acima): (1) um número mais elevado de eventos reprodutivos (e, portanto, descendentes) por vida se a mortalidade extrínseca dos adultos for baixa, (2) tempo suficiente para compensar a perda de descendentes devido à elevada mortalidade juvenil, e (3) menor incerteza reprodutiva devido à elevada variação da mortalidade juvenil de uma ronda de reprodução para a seguinte, mais uma vez podendo compensar a perda de descendentes. Estes efeitos são contrariados por aqueles que aumentam a mortalidade adulta (por exemplo, custos de sobrevivência da reprodução; envelhecimento) em relação à mortalidade juvenil. Assim, os aumentos na média (e/ou variância) da mortalidade adulta em relação à mortalidade juvenil tendem a favorecer uma vida reprodutiva mais curta e uma semelparidade, enquanto que as diminuições na média (e/ou variância) da mortalidade adulta em relação à mortalidade juvenil tendem a favorecer uma vida reprodutiva mais longa e uma iteroparidade.
Hoje em dia muitas destas previsões da teoria da história de vida são bem apoiadas por provas; terminaremos este artigo dando um exemplo de uma experiência particularmente elegante que confirmou as principais previsões do “problema geral da história de vida”.
Um exemplo de um Teste Empírico da Teoria
Para testar os princípios básicos da teoria da história de vida, Stearns et al. (2000) utilizaram uma população de moscas da fruta (D. melanogaster) para estabelecer dois conjuntos de linhas replicadas de “evolução experimental”: três linhas foram expostas a um tratamento de elevada mortalidade de adultos (HAM; matando aleatoriamente 90% das moscas duas vezes por semana) e três linhas a um tratamento de baixa mortalidade de adultos (LAM; matando 10% das moscas duas vezes por semana). Após quatro anos de evolução experimental nestas condições em laboratório, estelas e colaboradores mediram os fenótipos da história de vida de todas as linhas em ambos os tratamentos para examinar as respostas evolutivas das moscas à mortalidade extrínseca alta versus baixa. Os resultados desta experiência a longo prazo foram claros e intrigantes, confirmando as previsões teóricas: as moscas da fruta que tinham evoluído sob condições HAM desenvolveram-se mais rapidamente como larvas, fecharam mais cedo e com um tamanho mais pequeno como adultos, tiveram uma fecundidade precoce mais elevada, e mostraram uma duração de vida mais curta do que as moscas LAM que desenvolveram o conjunto oposto de adaptações. Por outras palavras, as moscas evoluindo num ambiente altamente perigoso responderam evolutivamente acelerando o seu desenvolvimento, diminuindo a sua idade e tamanho na maturidade, pondo mais ovos mais cedo, e vivendo menos tempo: adaptaram-se a níveis elevados de mortalidade aleatória, deslocando o seu esforço reprodutivo para idades mais precoces e comprimindo toda a sua história de vida para uma duração de vida mais curta. Assim, semelhantes aos resultados de Reznick et al. (1990) em guppies, estes resultados confirmam as principais previsões da teoria da história de vida, em particular o papel principal da mortalidade de adultos extrínsecos na moldagem da evolução do crescimento, maturação, reprodução, e envelhecimento.
Sumário
Aqui introduzimos as bases da teoria da história de vida. A teoria da história de vida tenta compreender como a selecção natural projecta os organismos para alcançar o sucesso reprodutivo, dado o conhecimento de como factores selectivos no ambiente (i.e., mortalidade extrínseca) e factores intrínsecos ao organismo (i.e., trade-offs, restrições) afectam a sobrevivência e a reprodução. Ao utilizar uma variedade de métodos teóricos e empíricos, e em particular ao aplicar o pensamento de optimização, os teóricos da história de vida derivaram grandes previsões sobre a evolução dos principais traços da história de vida, incluindo a idade e tamanho na maturidade, o número e tamanho dos descendentes, o esforço reprodutivo específico da idade ou tamanho, a duração da vida reprodutiva, e o envelhecimento. Com base nestas previsões, e testando-as em experiências de campo e laboratoriais, por exemplo em moscas, peixes, ou aves, os biólogos da história de vida forneceram-nos algumas respostas convincentes a questões fundamentais como, por exemplo, a idade e o tamanho: Com que rapidez deve um organismo desenvolver-se? Com que idade e tamanho deve amadurecer? Quantos descendentes deve ter e qual deve ser o seu tamanho? Deveria reproduzir-se uma ou mais vezes? E quanto tempo deve viver? Ao abordar estes problemas, a teoria da história de vida teve um grande impacto na nossa compreensão da adaptação por selecção natural, a questão mais fundamental em toda a biologia evolutiva.