古くからある農学的手法である接ぎ木は、植物の植物的・非植物的な繁殖の一種です。 通常、植物の損傷後のカルスの成長を利用して行われます。 接ぎ木は、園芸の分野では今でも一般的な技術です。 接ぎ木は、> 中国の紀元前2,000年頃から農業に用いられてきました。 農業、森林樹木の栽培、園芸植物の栽培などに広く利用されています。 例えば、経済的価値の高い地上部の品種の生育活力を向上させるために、病原菌に強い品種やストレス耐性のある品種を台木として使用するのが一般的です。 また、木本植物の開花を促進するために、幼木の段階を打破するためにも接ぎ木が用いられている。 遺伝子組み換え作物に比べて、接ぎ木された植物は一般の人にも受け入れられやすいものです。 接ぎ木は、植物科学の研究においても有用な技術です。 前世紀の70年代から80年代にかけて、この分野ではかなりの努力がなされてきました。 しかし、バイオテクノロジーの急速な発展に伴い、接ぎ木の研究の焦点は、農学的形質の改善から分子メカニズムの研究へと徐々に移行しています。
接ぎ木は、開花、オーキシン調節、重金属耐性、低分子RNAの動き、栄養状態、核サイレンシングなど、植物内の長距離シグナル伝達に関わるメカニズムを解明するための優れたツールであることが実証されています。
接ぎ木の生存率に影響を与える要因は、主に台木と穂木の相性に依存することがよく知られています。 互換性とは、台木と穂木の構造的、遺伝的、生理的特性に関する類似性と、接ぎ木後にお互いに適応する能力のことです。 同じ植物からの接ぎ木の相性が最も強く、植物同士の関係が遠いほど相性は弱くなります。 異なる属の植物を接ぎ木することは困難である。 穂木と台木の相性のほかに、台木と穂木の樹勢も接ぎ木の生存率に影響する重要な要素です。 また、接ぎ木された植物の生存率は種によって異なり、同じ種でも年齢が違えば接ぎ木体の生存率は異なる。 さらに、温度、湿度、光量などの環境要因も、接ぎ木植物の生存率に影響を与えます。
Arabidopsis thalianaとEutrema salsugineum(Thellungiella salsuginea)は同じCruciferae科に属しますが、異なる属です。 シロイヌナズナは双子葉植物の遺伝子研究のモデル植物として使われている。 Eutrema Salsugineum(別名:ソルトクレス)は、優れた耐塩性を持つ植物で、そのゲノムが完成している。 E. salsugineumの遺伝子の約95%はシロイヌナズナで発現している遺伝子と類似しており、アミノ酸配列もほとんどがシロイヌナズナのものと同じである。 Eutrema salsugineumの核ゲノムは比較的小さく、A. thalianaの約2倍の大きさである。 E. salsugineumのすべてのエコタイプは、寒冷、乾燥、酸化ストレスなどのさまざまな環境ストレスに抵抗性を示す。 E. salsugineumは、その極めて高い耐塩性により注目されている。 そのため、ソルトクレスは塩耐性を研究するためのモデルとなる塩生植物として20年以上前に提案された。 これにより、E. salsugineumをモデル植物として、環境ストレスだけでなく、光合成、ストレス関連タンパク質の機能、表面ワックスの生成など、幅広い分野の研究に新たな窓が開かれました。 シロイヌナズナでは、系統的なシグナル伝達を調べるために、いくつかの接ぎ木アプローチが開発されている。 しかし、我々の知る限り、E. salsugineumの接ぎ木方法や、シロイヌナズナとEutremaの接ぎ木方法についての報告はありません。 これらの植物間の接ぎ木の難易度は著しく高い。 この2つのモデル植物の接ぎ木に成功すれば、植物のストレス耐性の分子メカニズムに関する重要な洞察を得ることができます。
接ぎ木の方法
本研究では、塩分に弱いシロイヌナズナと塩分に強いソルトクレスの間で、2つの効率的な無菌苗の接ぎ木方法に注目しました。 1つの方法は、SeungおよびTurnbullらによって記述されているように、苗の胚軸-下胚軸の平面切断であり、いくつかの変更が加えられている。 もう1つの方法は、本明細書で実証するカットイン接ぎ木であった。 ここで使用される略語A/Eは、A. thalianaの穂木がE. salsugineumの株に接ぎ木されたことを示し、逆にE/Aは、E. salsugineumの穂木がA. thalianaの株に接ぎ木されたことを示す。 対照実験として、A. thalianaとE. salsugineumの自己接ぎ木を行った。 A. thalianaの自己接ぎ木苗をA/A、E. salsugineumの自己接ぎ木苗をE/Eと呼ぶ。 最適化された接ぎ木方法が、異なる台木と穂木に適用された。 A/Eにはカットイン接ぎ木の方法を用い(図1a)、A/A、E/E、E/Aにはその他の方法、すなわち胚軸-下胚軸の平板切断を用いた(図1c、d)。 従来の接ぎ木方法(胚軸-下胚軸の平板切断)では、接ぎ木を成功させることは比較的容易であり、その後の接ぎ木植物の生育も通常は満足できるものである。

接ぎ木技術の模式図 a, b Eutrema.Salsugineumに接ぎ木したArabidopsis thalianaの2つの異なる接ぎ木方法。 Salsugineum(A/E)、切り込み接ぎ(a)と胚軸-下胚軸の平面切り(b)。 c E/A(E. salsugineumを穂木としてA.thalianaにストックとして接ぎ木した。 d)E. salsugineum(E/E)とA. thaliana(A/A)の自己接ぎ木。 赤い線は切断位置を示す。 オレンジ色の矢頭は、シリコンチューブ支持体(ライトグレー)を用いたシュートと根の組織間の接ぎ木接合部を示す。 (a)の赤矢印は切断・除去部分を示す。 色の違いは異なる植物を示しており、E. salsugineumは青、A. thalianaは緑である。 根はここでは示していない
材料と生育条件
本研究で使用したArabidopsis thaliana Columbia-0 (Col)の種子とE. salsugineum (Shandong, China)の種子は、当研究室で保存・増殖したものである。
接ぎ木の手順
A/Eに適用したカットイン接ぎ木法のステップバイステップのプロトコルは以下の通りです:
台木と穂木の材料の準備:ソルトクレスの種子を0.5%のNaClO(塩酸ナトリウム)で6分間消毒した後、蒸留水で4-5回洗浄し、無菌状態で、1.0%の寒天と1%の砂糖を含む半分の強度のムラシゲ・スクーグ(MS)培地に播種した。 低温(4℃)成育処理の1週間後にSD条件に移し、10-15日間(または発芽後7-10日間)光を当てて垂直培養し、根株苗とした。 シロイヌナズナ種子の発芽については、低温(4℃)成層処理が2~3日であったことを除き、殺菌方法および生育条件はE. salsugineumと同様であった。 シロイヌナズナの苗を接ぎ木するのに最適な年齢は3~5日である。 2つの種の発芽時間と成長速度が異なるため、それに応じて植え付け時期を調整した。 通常、シロイヌナズナはソルトクレスの種子が発芽した後に植えるべきである。
-
無菌状態での接ぎ木(図1a)。 接ぎ木の手順は、無菌フードの中で解剖用顕微鏡の下で行われた。 均一な大きさのE. salsugineumの苗をピンセットで新しい1.0%半固形MS寒天培地に移した。 根株として使用するために、ソルトクレスの苗からステンレス製の両刃のカミソリで本葉と分裂組織を取り除き、子葉は残した。 同じ無菌状態で、A. thalianaの苗の胚軸を上から高さの約1/4(通常2〜3mm)で横に切り、苗の上部を穂木として使用した。 穂木としてのA.thalianaの切断は、元の培地プレートで直接行うこともできるし、ソルトクレスを根株として調製したプレートで行うこともできる。 穂木のソルトクレスへの接ぎ木は、2つの部分の切断面にある微量の液体で十分に保持されるため、固定具を使わずに行った。 その後、湿度を保つためにパラフィルムでペトリ皿を素早く、しかし丁寧に密封しました。 なお、植物の成長により葉が外れる可能性がありますので、葉が自然に伸びるように注意してください。
-
移植体の培養。 接ぎ木されたA/E複合体は、パーシバル社のグロースキャビネットでSD条件で6~8日間置いた。
-
接ぎ木苗の移植(図2)。
-
移植(Fig.2):移植後6〜8日目に、解剖学的顕微鏡でバットジョイントの界面を観察した。 うまくいった接ぎ木は、この時までに穂木と株の間がしっかりとつながっているはずである。 その後、苗を必要に応じて水耕栽培システムや栄養価の高い土壌に移植した。
Fig.2のように、不定根が発生した場合には、適時、不定根を切断するように注意します。 2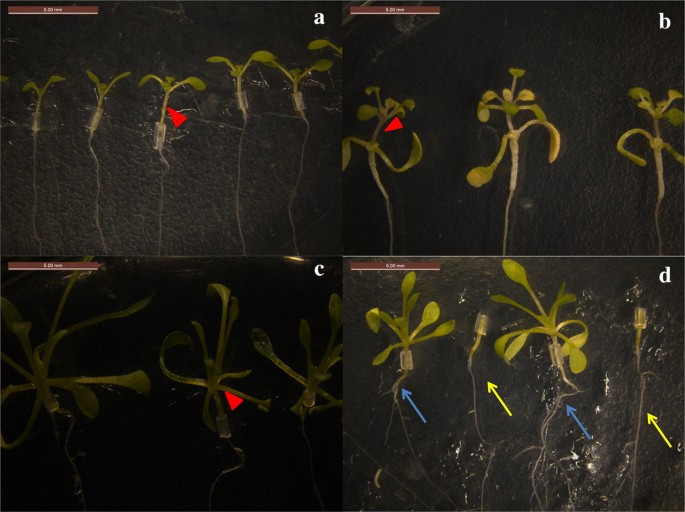
成功した接ぎ木苗。a A.thalianaの自殖接ぎ木(A/A)。b A/Eの接ぎ木苗。c E.salsugineumの自殖接ぎ木(E/E)。d E/Aの接ぎ木苗。 a-cの赤矢印は接ぎ木部位を示す。 dではシロイヌナズナの根を異なる色の矢印で示し、青矢印は接ぎ木後の根を示す。
種子表面の消毒
カットイングラフト法では、種子表面を0.5%塩酸ナトリウム(NaClO)で6-8分間直接消毒し、その後、滅菌水で4-6回洗浄した。 この方法では、アルコールによる殺菌を避けることができます。 また、NaClOによる殺菌方法は簡便であるため、二次汚染が少なく、種子は正常に発芽します。 また、殺菌しすぎて種子が発芽しなかったり、その後の分析に影響が出たりすることも避けられます。
苗代
カットイン接ぎ木法では、やや古いE. salsugineumの苗(発芽後7-10日目)に側根を付けておくと、穂木(A. thaliana)の早い成長の特性をよりよく満たすことができ、また不定根の形成を抑え、接ぎ木の生存率を大幅に向上させ、後の段階での接ぎ木苗の旺盛な成長を保証することができます(表1)。 その結果、シロイヌナズナの3〜5日齢の苗は、接ぎ木苗の生存率を向上させるのに有利であり、また、接ぎ木苗の旺盛な成長をサポートすることがわかりました(表1)。 これは、シロイヌナズナのライフサイクルが、E. salsugineumに比べて短いためと考えられる。 そのため、古いA. thalianaの苗を穂木として使用すると、生存率が低くなり、A/Eグラフトが生き残ったとしても、その後の生育が悪く、通常は早期に開花することが多いのです。
表1 A/E接ぎ木における苗の年齢の変化 平板切断法
A. thalianaでは、複数の種類の接ぎ木法が報告されている。 本研究では、A/E接ぎ木には切り込み接ぎ木の方法のみを用い、A/A、E/E、E/Aには胚軸-下胚軸の平面切断の方法を用いた(図1c、d)。 この2つの方法は,切断部位を除いて同じ培養条件を必要とする。 A/Aセルフグラフトには、3~5日齢の苗を用いた。 E/A接ぎ木では、A. thalianaの7~10日齢の苗を台木として用いた方が、接ぎ木の生存率を高めることができる。 同様に、E/Eセルフグラフトには、7~10日齢の苗を使用した。 その結果、2つの接ぎ木方法の間で、A/E苗の生存率とその後の生長に有意な差があることがわかった(図1a、bおよび表2)。 A/Eの接ぎ木では、切り込み法の方が満足度が高い。 その主な理由は、A. thalianaの速い成長速度が、この方法を用いてよりよく対応できるためであると考えられる。 A/E接ぎ木に従来の平面カットを用いた場合、ソルトクレスはA. thalianaよりも成長が遅いため、接合部のカルス形成前に穂木からの不定根が現れ、接ぎ木の成功率に影響を与えた。
表2 2つの接ぎ木方法のA/E接ぎ木への影響 。
