Variatie van de bloedsomloop bij dieren
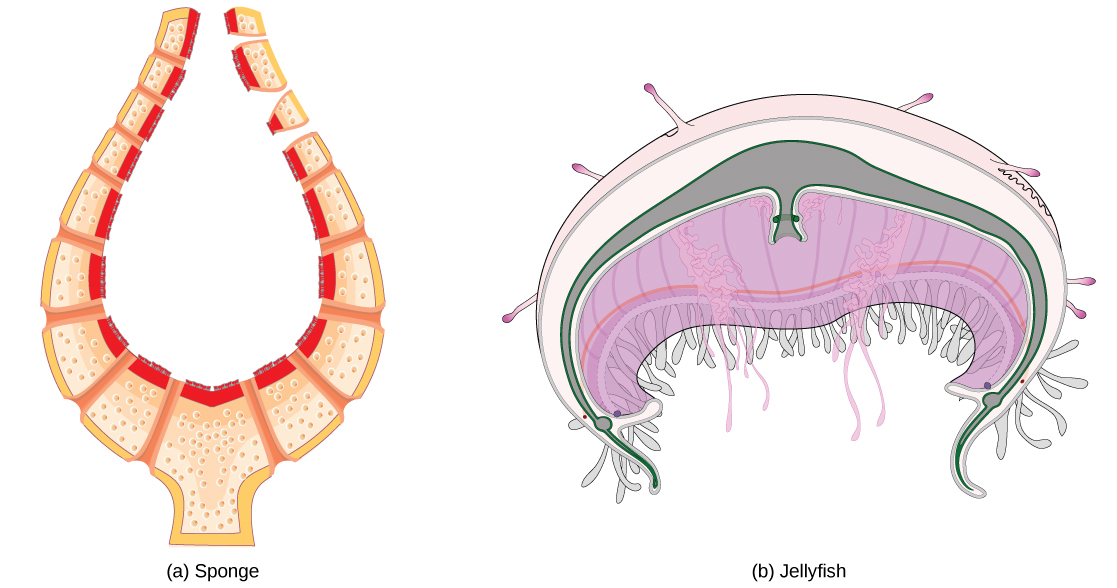
De bloedsomloop varieert van eenvoudige systemen bij ongewervelde dieren tot meer complexe systemen bij gewervelde dieren. De eenvoudigste dieren, zoals de sponzen (Porifera) en de rotiferen (Rotifera), hebben geen bloedsomloop nodig omdat diffusie zorgt voor een adequate uitwisseling van water, voedingsstoffen en afval, alsmede opgeloste gassen, zoals weergegeven in (Figuur)a. Organismen die complexer zijn maar nog steeds slechts twee cellagen in hun lichaamsplan hebben, zoals jellies (Cnidaria) en kammellies (Ctenophora), maken ook gebruik van diffusie via hun opperhuid en inwendig via het gastrovasculaire compartiment. Zowel hun inwendige als hun uitwendige weefsels baden in een waterig milieu en wisselen vloeistoffen uit door diffusie aan beide zijden, zoals geïllustreerd in (figuur) b. De uitwisseling van vloeistoffen wordt bevorderd door het pulseren van het kwallenlichaam.
Voor complexere organismen is diffusie niet efficiënt om gassen, voedingsstoffen en afvalstoffen effectief door het lichaam te laten circuleren; daarom zijn complexere bloedsomloopsystemen geëvolueerd. De meeste geleedpotigen en veel weekdieren hebben een open bloedsomloop. In een open systeem duwt een langgerekt kloppend hart de hemolymfe door het lichaam en helpen spiersamentrekkingen om de vloeistoffen te verplaatsen. De grotere, meer complexe schaaldieren, waaronder kreeften, hebben arteriële vaten ontwikkeld om het bloed door hun lichaam te persen, en de meest actieve weekdieren, zoals inktvissen, hebben een gesloten bloedsomloop ontwikkeld en zijn in staat zich snel te verplaatsen om prooien te vangen. De gesloten bloedsomloop is een kenmerk van de gewervelde dieren; er zijn echter aanzienlijke verschillen in de structuur van het hart en de bloedsomloop tussen de verschillende groepen gewervelde dieren als gevolg van aanpassingen tijdens de evolutie en daarmee samenhangende verschillen in anatomie. (Figuur) illustreert de basisbloedsomloop van enkele gewervelde dieren: vissen, amfibieën, reptielen en zoogdieren.
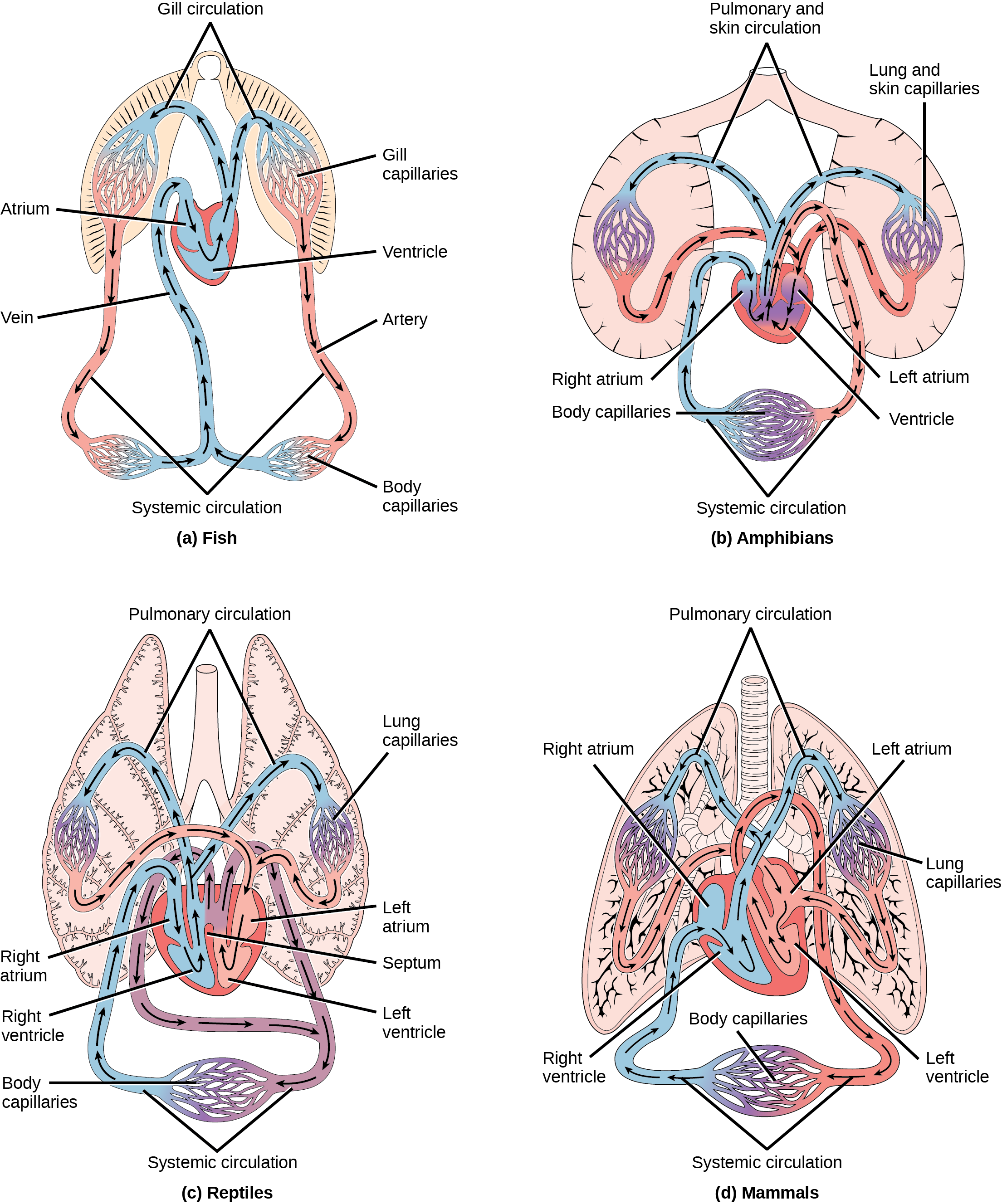
Zoals geïllustreerd in (figuur)a. Vissen hebben een enkel circuit voor de bloedstroom en een tweekamerhart dat slechts één atrium en één ventrikel heeft. Het atrium vangt het bloed op dat uit het lichaam is teruggekeerd en de ventrikel pompt het bloed naar de kieuwen waar gasuitwisseling plaatsvindt en het bloed opnieuw zuurstof krijgt; dit wordt kieuwcirculatie genoemd. Het bloed stroomt vervolgens door de rest van het lichaam voordat het weer bij het atrium aankomt; dit wordt de systemische circulatie genoemd. Deze eenrichtingsstroom van bloed veroorzaakt een gradiënt van zuurstofrijk naar zuurstofarm bloed rond het systemische circuit van de vis. Het resultaat is een beperking van de hoeveelheid zuurstof die sommige organen en weefsels van het lichaam kan bereiken, waardoor de totale stofwisselingscapaciteit van vissen afneemt.
Bij amfibieën, reptielen, vogels en zoogdieren verloopt de bloedstroom in twee circuits: een door de longen en terug naar het hart, die pulmonale circulatie wordt genoemd, en de andere door de rest van het lichaam en de organen, waaronder de hersenen (systemische circulatie). Bij amfibieën vindt de gasuitwisseling ook plaats via de huid tijdens de pulmonale circulatie; dit wordt pulmocutane circulatie genoemd.
Zoals te zien is in (figuur)b, hebben amfibieën een driekamerhart met twee atria en één ventrikel, in plaats van het tweekamerhart van vissen. De twee boezems (superieure hartkamers) ontvangen bloed uit de twee verschillende circuits (de longen en de stelsels), en vervolgens is er enige vermenging van het bloed in de ventrikel (inferieure hartkamer) van het hart, wat de efficiëntie van de oxygenatie vermindert. Het voordeel van deze opstelling is dat de hoge druk in de vaten het bloed naar de longen en het lichaam stuwt. De vermenging wordt afgezwakt door een ribbel in de ventrikel die zuurstofrijk bloed afvoert via de systemische bloedsomloop en zuurstofarm bloed naar het pulmocutane circuit. Om deze reden wordt van amfibieën vaak gezegd dat zij een dubbele circulatie hebben.
De meeste reptielen hebben ook een driekamerhart, vergelijkbaar met het amfibieënhart, dat het bloed naar de pulmonale en systemische circuits leidt, zoals te zien is in (figuur)c. De ventrikel wordt doeltreffender verdeeld door een gedeeltelijk septum, waardoor minder vermenging van zuurstofrijk en zuurstofarm bloed optreedt. Sommige reptielen (alligators en krokodillen) zijn de primitiefste dieren die een vierkamerhart vertonen. Krokodilachtigen hebben een uniek mechanisme van bloedsomloop waarbij het hart het bloed van de longen naar de maag en andere organen leidt gedurende lange perioden van onderdompeling, bijvoorbeeld terwijl het dier wacht op een prooi of onder water blijft wachten tot de prooi verrot is. Eén aanpassing omvat twee hoofdslagaders die uit hetzelfde deel van het hart komen: de ene voert het bloed naar de longen en de andere zorgt voor een alternatieve route naar de maag en andere delen van het lichaam. Twee andere aanpassingen zijn een gat in het hart tussen de twee hartkamers, het zogenaamde foramen van Panizza, waardoor bloed van de ene kant van het hart naar de andere kan stromen, en gespecialiseerd bindweefsel dat de bloedstroom naar de longen vertraagt. Samen hebben deze aanpassingen krokodillen en alligators tot een van de meest evolutionair succesvolle diergroepen op aarde gemaakt.
Bij zoogdieren en vogels is het hart ook verdeeld in vier kamers: twee boezems en twee kamers, zoals geïllustreerd in (figuur)d. Het zuurstofhoudende bloed wordt gescheiden van het zuurstofarme bloed, hetgeen de doelmatigheid van de dubbele circulatie verbetert en waarschijnlijk noodzakelijk is voor de warmbloedige levenswijze van zoogdieren en vogels. Het vierkamerhart van vogels en zoogdieren evolueerde onafhankelijk van een driekamerhart. De onafhankelijke evolutie van dezelfde of een soortgelijke biologische eigenschap wordt convergente evolutie genoemd.